ARTICLE INFORMATION
Copyright statement: ? XXX
Copyright: 2013
Date received: 25 June 2012
Date accepted: 17 January 2013
Publication date (electronic): November 2013
Volume: 4
Issue: 1
Pages: 1-11
Publisher ID: xxxxxx
Epub and EBook conversion are additional services for publishers, researchers and editors
All Epubs are validated from International Digital Publishing Forum idfp.
In specialist evolving, JATS TECH take great care in the conversion process, to produce eBooks that are pleasant to look at, easy to read and follow the layout of the original printed book, as closely as possible.
JATS TECH will provide for review in both Amazon MOBI and EPUB formats.
EPUB 2.0.1 is a maintenance release of EPUB 2. Its development was chartered in 2009, and the final standard was approved by the IDPF Membership as a Recommended Specification in May, 2010.
EPUB 2.0.1 is defined by three open standard specifications, the Open Publication Structure (OPS), Open Packaging Format (OPF) and Open Container Format (OCF).
EPUB 3 was developed pursuant to a charter approved by the IDPF Membership in May, 2010. EPUB 3.0 was approved by the IDPF membership as a final Recommended Specification effective October 11, 2011, and superseded the previous release of EPUB (2.0.1).
This page contains information about EPUB 3.0 which as of June, 2014 has been superseded by EPUB 3.0.1, which is now the current version of the EPUB standard.
All production of EPUBs is validated from International digital publishing forum. Below is the working of JATS TECH digital publishing engineers. Click the button for viewing







Journal ID (publisher-id): XXX
Title: XXXX
Abbreviated Title: XXXX
ISSN (electronic): 1234-5678
Publisher: XXXXX
Copyright statement: ? XXX
Copyright: 2013
Date received: 25 June 2012
Date accepted: 17 January 2013
Publication date (electronic): November 2013
Volume: 4
Issue: 1
Pages: 1-11
Publisher ID: xxxxxx
Aijaz Ahmad Wani*, Shahzada Arshid
Department of Botany, University of Kashmir, Kashmir, India
* Corresponding author, aijazbotku@gmail.com
The present study was undertaken to understand the fruit and seed morphology, seed viability and effect of various physical and chemical factors on seed germination allowing us to explore the spread potential and/or seedling recruitment mechanism in Myriophyllum spicatum L.. The fruit of the species is a schizocarp, while as seed is a nutlet. The seed set was recorded to be ranging from 70.98-77.91% across the standing water populations, whereas no seed set was observed in running water populations due to the lack of an effective pollination system. The seed viability ranged from 85-90%. For in-vitro seed germination studies, the seeds were subjected to different physical and chemical treatments under alternate light and dark as well as continuous dark conditions. The seeds in control and those treated with different concentrations of GA3 and IAA and those whose epicarp and mesocarp were removed did not show any signs of germination. However, it was observed that surgical exposure of the embryo (cutting of hard endocarp of seed) has a promoter effect on germination and maximum percentage germination (76.66 ± 5.77) was recorded due to surgical exposure of embryo plus different concentrations of GA3. Moreover, a good germination percentage was recorded in seeds subjected to chilling treatment. Further, it was observed that seed germination of one-year-old seeds was less if compared to the current year seeds and overall percentage germination was higher in alternate light and dark if compared to continuous dark conditions in all the treatments.Thus, we conclude that the dormancy of the seeds is due to the hard endocarp and that the light has a promoting effect on germination. With the increase in the age of the seeds, there is decrease in their viability and hence germination. The chilling winter temperature of the Kashmir is responsible for breaking the hard endocarp of the seeds leading to their germination and hence spread of the populations.
Keyword: seed dormancy, light, chilling, GA3, indoleacetic acid
O presente estudo foi realizado para compreender a morfologia do fruto e da semente, a viabilidade da semente e o efeito de vãrios fatores físicos e químicos na germinação de sementes, permitindo-nos explorar o potencial de disseminação e/ou mecanismo de mudas de recrutamento em Myriophyllum spicatum L. O fruto da espécie é um chisocarpo, enquanto a semente é uma noz. A produção de sementes variou de 70,98 a 77,91% entre as populações de ãgua paradas, enquanto que nenhum conjunto de sementes foi observado em populações de ãgua corrente, devido à falta de um sistema eficaz de polinização. A viabilidade das sementes variou de 85-90%. Para estudos in vitro de germinação das sementes, as sementes foram submetidas a diferentes tratamentos físicos e químicos sob luz e escuridão alternadas, bem como contínuo escuro. As sementes de controle e aquelas tratados com diferentes concentrações de GA3 e IAA e aquelas cujo epicarpo e mesocarpo foram removidos não mostraram quaisquer sinais de germinação. No entanto, observou-se que a exposição cirúrgica do embrião (corte do endocarpo rígido de semente) tem um efeito sobre a germinação e efeito promotor sobre a germinação e um percentual mãximo (76,66 ± 5,77) foi registada por exposição cirúrgica do embrião a diferentes concentrações de GA3. Além disso, a percentagem de germinação foi observada em sementes submetidas ao tratamento de resfriamento. Ademais, observou-se que a germinação de sementes de um ano de idade foi menor em comparação com as sementes do ano corrente e a percentagem de germinação em geral foi maior quando houve luz e escuridão alternadasse comparadas com as em condições de escuridão em todos os tratamentos. Assim, podese concluir que a dormência das sementes é atribuída à rigidez de endocarpo e que a luz tem um efeito promotor sobre a germinação. Com o aumento da idade das sementes, hã redução na viabilidade e, portanto, na germinação destas. A temperatura do inverno frio da Caxemira é responsãvel por quebrar o endocarpo duro das sementes, o que leva à sua germinação e, consequentemente, à disseminação das populações.
Palavras-chave: dormência de sementes, GA3, luz, refrigeração, ãcido idol acético
Myriophyllum spicatum L. commonly known as watermilfoil is a rooted perennial, widely distributed in Kashmir Himalaya and can be found in almost all kinds of freshwater habitats including lakes, ponds, irrigation ditches and rivers (Arshid et al., 2011). It is native to Europe, Asia and North America (Couch & Nelson, 1985). From the initial point of introduction in the North America, M. spicatum has spread to 46 states and three Canadian provinces of North America (Jacono & Richerson, 2003). It is categorized among the five most ubiquitous, noxious and the most widely managed aquatic weed in lake ecosystems (Bartodziej &Ludlow, 1998; Marsden & Hauser, 2009).The importance of seeds in the propagation of Myriophyllum spicatum may lie in their ability to remain viable following various disturbances serving as an effective long-term mechanism of survival and regrowth.
Although seed germination of terrestrial species has been extensively studied, few studies have examined submerged macrophytes (Baskin & Baskin, 1988; Hartleb et al., 1993; Lal & Gopal, 1993; Ke & Li, 2006; Hay et al., 2008; Jarvis & Moore, 2008). This may be due to the fact that most submerged macrophytes show dominant clonal reproduction, and their seeds are rarely observed to grow into mature plants in nature. However, seeds of submerged macrophytes have been found to play important roles in the establishment of new genotypes in existing populations (Kimber et al., 1995), movement of populations into new regions (Arnold et al., 2000; Figuerola & Green, 2002; Harwell & Orth, 2002) and re-establishment of populations after episodic declines (McMillan & Jewett-Smith, 1988; Titus & Hoover, 1991; Parveen et al., 1995; Jarvis & Moore, 2008). A recent study on the genetic pattern of Potamogeton malaianus has suggested that bird mediated seed dispersal could greatly influence gene movements among lakes (Chen et al., 2009). Germination requirements and dormancy characteristics of species are often assumed to be adaptations to the particular habitats in which the species occur (Angevine & Chabot, 1979; Meyer et al., 1990). Germination at the right time and in the right place is important to determine the probability of a seedling surviving to maturity (Thompson, 1973). Previous studies have indicated that temperature (Hartleb et al., 1993; Ke & Li, 2006; Hay et al., 2008; Jarvis & Moore, 2008) and light (Coble & Vance, 1987; Lal & Gopal, 1993) are important ecological triggers in the seed germination of submerged species. Xiao et al. (2010) have examined the effect of temperature, water level and burial depth on seed germination of Myriophyllum spicatum and Potamogeton malaianus. Madsen & Boylen (1989),while studying the seed germination of M. spicatum from an oligotrophic and eutrophic lake, have concluded that in oligotrophic lake, the seeds do not contribute to the spread of the population due to the low seed set and low rate of germination, while in eutrophic lake there was a potential for seeds to contribute to annual propagation and expansion of the population due to relatively high seed set percentages and germination rates. In this background, understanding the seed set, viability, and germination characteristics of M.spicatum may reveal a mechanism for species invasion.
In the present study, we have studied the fruit and seed morphology, seed set, seed viability and quantified the effect of different physical as well as chemical treatments on percentage seed germination what have allowed us to explore the spread potential and/or seedling recruitment mechanism in the species.
In the present study, fresh and one year old seeds of Myriphyllum spicatum L. were collected in October, 2010,from three standing water sites namely Dal Lake (DL), Mansbal Lake (ML) and Hygam Wetland (HW). The two running water sites, namely Shalimar Stream (SS) and Chanderhama Irrigation Canal (CIC), did not produce any seed.One part of the seed lot was used one year later in 2011 for germination studies. The seed quality parameters were studied along the following lines,
For estimating the seed set and reproductive success, plants were selected, at random, in different populations, tagged and scored for the number of spikes per ramet and the number of flowers and seeds per spike, following Lubber &Christensen (1966). Percentage seed set was calculated as follows:

Seed viability was analyzed using Tz-test. Twenty seeds of each replicate were placed in water, at room temperature, for 24 hours. The imbibed seeds were, then, longitudinally sectioned and the sections were incubated in dark in 2% aqueous solution of Tz, for 24 hours. The dissected seeds having healthy embryo and stain deep red in 2% 2,3,5 triphenyl tetrazolium chloride were considered as viable (Hendry & Grime, 1993).
For in vitro seed germination studies, freshly collected and one year old seeds from natural population were used. The seeds were washed with 0.1% mercuric chloride for 5-7 minutes, followed by washing 4-5 times with distilled water and subjected to different physical and chemical treatments (Table 1). All the experiments were conducted in temperature, light and humidity controlled incubation cabinets. The treatment solutions were made using deionized water and analytical grade chemicals. Each treatment consisted of three replicates of 10 seeds, placed within closed glass petri-dishes on Wattman filter paper moistened with 15 ml of the given treatment solution or distilled water. Seeds planed for dark treatment were moistened under green light and then quickly wrapped in black carbon papers to reduce the chances of exposure to light. Emergence of plumule was used as the indicator of germination and total germination was recorded at the culmination of the experiment in respect of all the treatments. Seed germination was also checked by placing 50 healthy and viable seeds in 30 x 10 x 4 inch tray containing 2 kg (wet mass) of sediment and the water levels were maintained 1.5 inch above the surface of the sediment. These trays were placed inside the seed germinator, where temperature was maintained at 25±2°c. Seeds were considered to have germinated at the first visible plumule emergence.
Physical and chemical treatments to test seed germination in Myriophyllum spicatum.
Scanning Electron Microscopic (SEM) Studies
For scanning electron microscopic studies, the whole seeds and fruits were loaded on the stub and then coated with gold and observed under SEM (S-3000H) at the University Scientific Instrumation Centre.
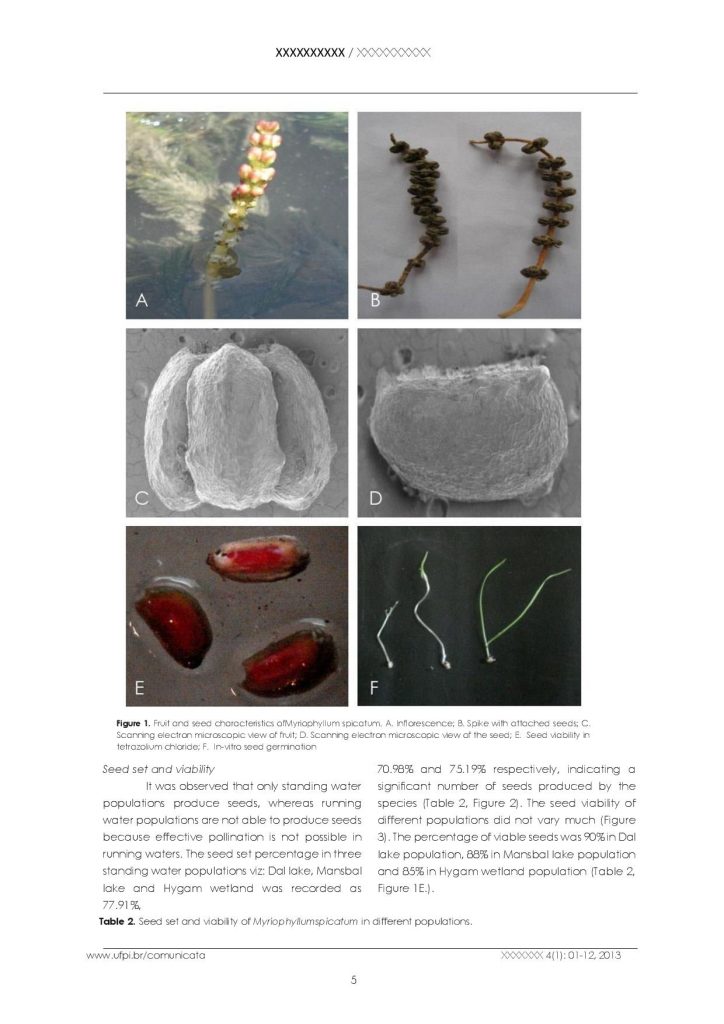
In Myriophyllum spicatum the inflorescence is a cylindrical spike raised on an elongated peduncle with acropetally arranged unisexual flowers.The upper flowers are staminate whereas lower ones are pistillate (Figures 1A, B).The scanning electron microscopic analysis revealed that fruit is a globular schizocarp measuring 2.2×2.5 mm having smooth surface architecture with four deep longitudinal sutures and a square shaped scar on upper side having four elongated points, which are the remains of the stigmas (Figure 1C). The lower side (from which fruit is attached to the spike) is circular. Each fruit contains four individual seeds. The seed is a trigonal nutlet 2.2×1.5 mm with smooth epicarp and mesocarp and a hard endocarp,having two flat sides and an outer convex side (Figure 1D).
Fruit and seed characteristics ofMyriophyllum spicatum. A. Inflorescence; B. Spike with attached seeds; C. Scanning electron microscopic view of fruit; D. Scanning electron microscopic view of the seed; E. Seed viability in tetrazolium chloride; F. In-vitro seed germination

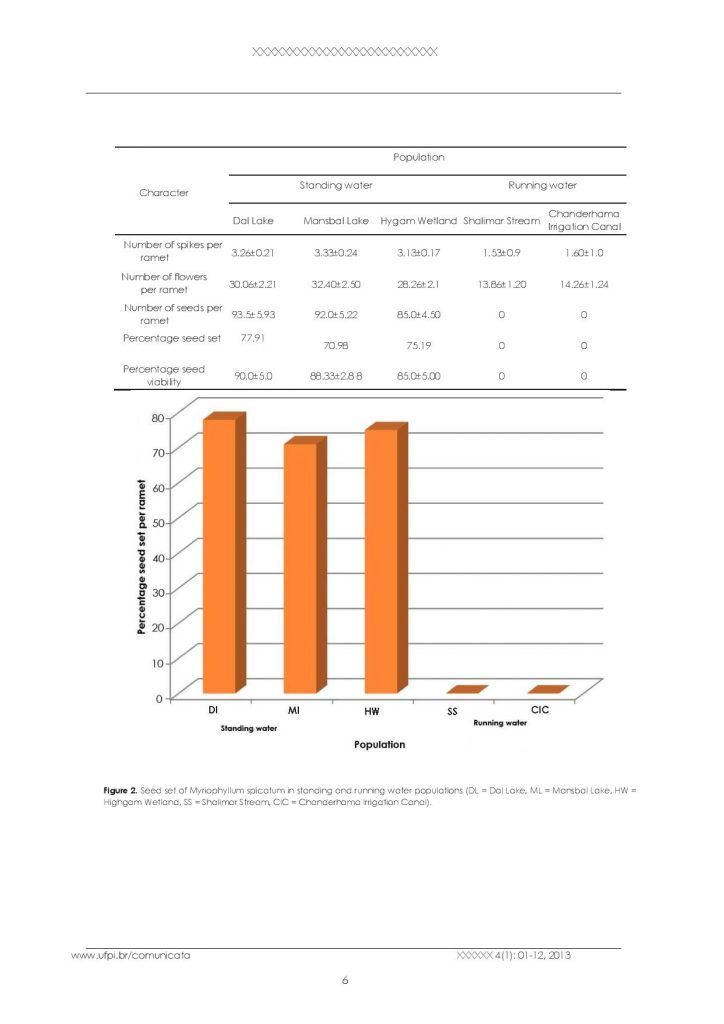
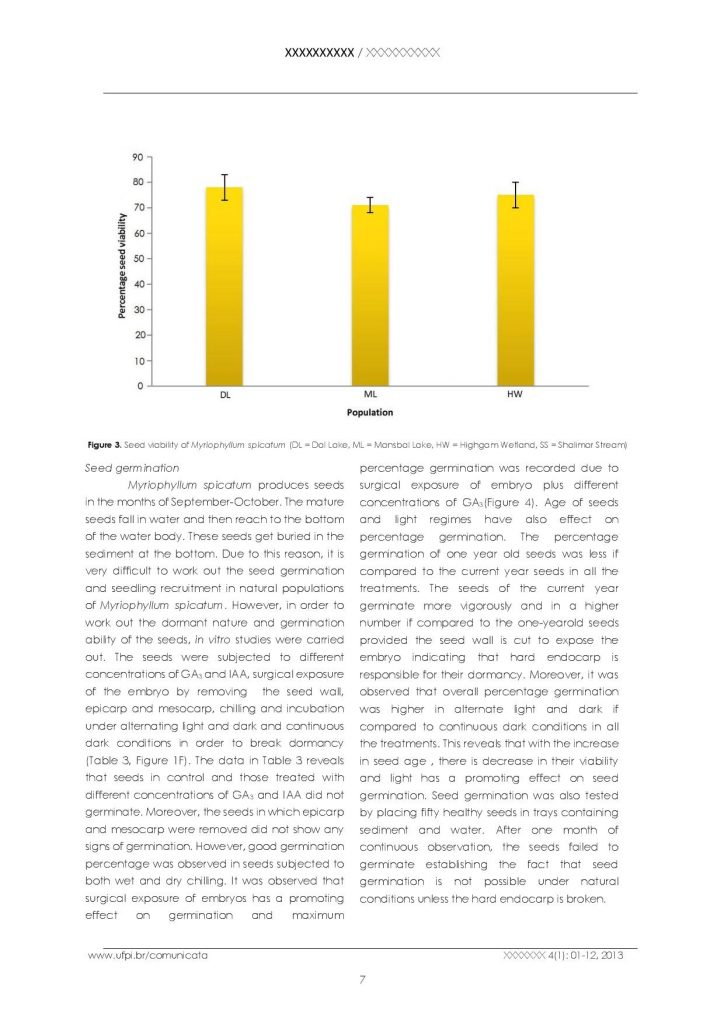
It was observed that only standing water populations produce seeds, whereas running water populations are not able to produce seeds because effective pollination is not possible in running waters. The seed set percentage in three standing water populations viz: Dal lake, Mansbal lake and Hygam wetland was recorded as 77.91%, 70.98% and 75.19% respectively, indicating a significant number of seeds produced by the species (Table 2, Figure 2). The seed viability of different populations did not vary much (Figure 3). The percentage of viable seeds was 90% in Dal lake population, 88% in Mansbal lake population and 85% in Hygam wetland population (Table 2, Figure 1E.).
Seed set and viability of Myriophyllumspicatum in different populations.
Seed set of Myriophyllum spicatum in standing and running water populations (DL = Dal Lake, ML = Mansbal Lake, HW = Highgam Wetland, SS = Shalimar Stream, CIC = Chanderhama Irrigation Canal).

Seed viability of Myriophyllum spicatum (DL = Dal Lake, ML = Mansbal Lake, HW = Highgam Wetland, SS = Shalimar Stream)

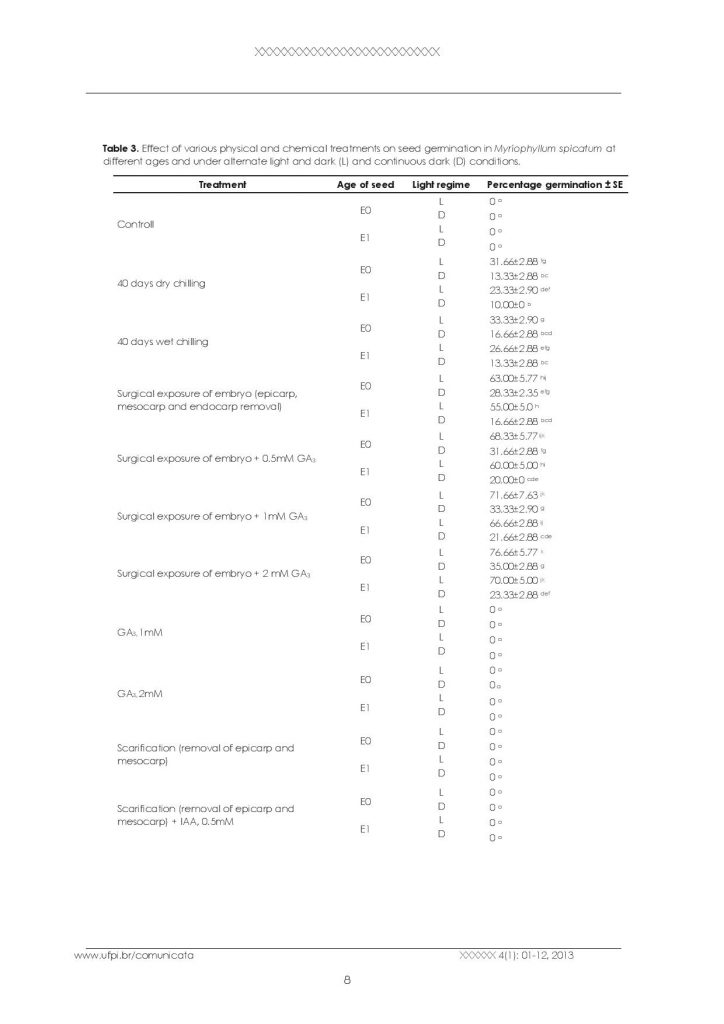
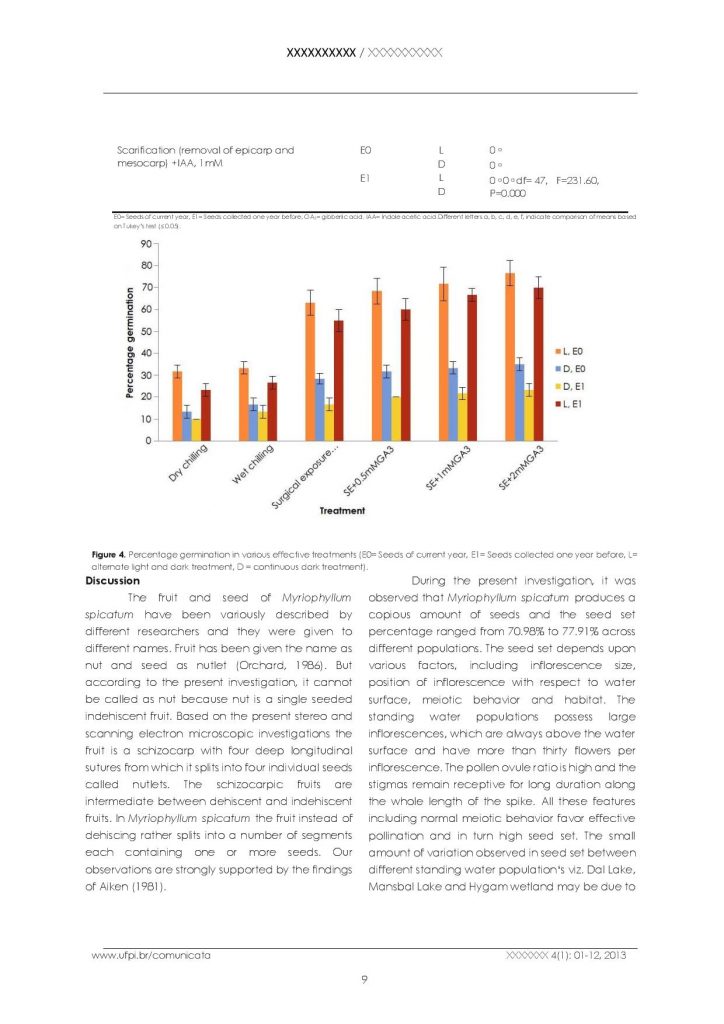
Myriophyllum spicatum produces seeds in the months of September-October. The mature seeds fall in water and then reach to the bottom of the water body. These seeds get buried in the sediment at the bottom. Due to this reason, it is very difficult to work out the seed germination and seedling recruitment in natural populations of Myriophyllum spicatum. However, in order to work out the dormant nature and germination ability of the seeds, in vitro studies were carried out. The seeds were subjected to different concentrations of GA3 and IAA, surgical exposure of the embryo by removing the seed wall, epicarp and mesocarp, chilling and incubation under alternating light and dark and continuous dark conditions in order to break dormancy (Table 3, Figure 1F). The data in Table 3 reveals that seeds in control and those treated with different concentrations of GA3 and IAA did not germinate. Moreover, the seeds in which epicarp and mesocarp were removed did not show any signs of germination. However, good germination percentage was observed in seeds subjected to both wet and dry chilling. It was observed that surgical exposure of embryos has a promoting effect on germination and maximum percentage germination was recorded due to surgical exposure of embryo plus different concentrations of GA3(Figure 4). Age of seeds and light regimes have also effect on percentage germination. The percentage germination of one year old seeds was less if compared to the current year seeds in all the treatments. The seeds of the current year germinate more vigorously and in a higher number if compared to the one-yearold seeds provided the seed wall is cut to expose the embryo indicating that hard endocarp is responsible for their dormancy. Moreover, it was observed that overall percentage germination was higher in alternate light and dark if compared to continuous dark conditions in all the treatments. This reveals that with the increase in seed age , there is decrease in their viability and light has a promoting effect on seed germination. Seed germination was also tested by placing fifty healthy seeds in trays containing sediment and water. After one month of continuous observation, the seeds failed to germinate establishing the fact that seed germination is not possible under natural conditions unless the hard endocarp is broken.
Effect of various physical and chemical treatments on seed germination in Myriophyllum spicatum at different ages and under alternate light and dark (L) and continuous dark (D) conditions.
| Treatment | Age of seed | Light regime | Percentage germination ± SE |
|---|---|---|---|
| Controll | E0 | L | 0 a |
| D | 0 a | ||
| E1 | L | 0 a | |
| D | 0 a | ||
| 40 days dry chilling | E0 | L | 31.66±2.88 fg |
| D | 13.33±2.88 bc | ||
| E1 | L | 23.33±2.90 def | |
| D | 10.00±0 b | ||
| 40 days wet chilling | E0 | L | 33.33±2.90 g |
| D | 16.66±2.88 bcd | ||
| E1 | L | 26.66±2.88 efg | |
| D | 13.33±2.88 bc | ||
| Surgical exposure of embryo (epicarp, mesocarp and endocarp removal) | E0 | L | 63.00±5.77 hij |
| D | 28.33±2.35 efg | ||
| E1 | L | 55.00±5.0 h | |
| D | 16.66±2.88 bcd | ||
| Surgical exposure of embryo + 0.5mM GA3 | E0 | L | 68.33±5.77 ijk |
| D | 31.66±2.88 fg | ||
| E1 | L | 60.00±5.00 hi | |
| D | 20.00±0 cde | ||
| Surgical exposure of embryo + 1mM GA3 | E0 | L | 71.66±7.63 jk |
| D | 33.33±2.90 g | ||
| E1 | L | 66.66±2.88 ij | |
| D | 21.66±2.88 cde | ||
| Surgical exposure of embryo + 2 mM GA3 | E0 | L | 76.66±5.77 k |
| D | 35.00±2.88 g | ||
| E1 | L | 70.00±5.00 jk | |
| D | 23.33±2.88 def | ||
| GA3, 1mM | E0 | L | 0 a |
| D | 0 a | ||
| E1 | L | 0 a | |
| D | 0 a | ||
| GA3, 2mM | E0 | L | 0 a |
| D | 0 a | ||
| E1 | L | 0 a | |
| D | 0 a | ||
| Scarification (removal of epicarp and mesocarp) | E0 | L | 0 a |
| D | 0 a | ||
| E1 | L | 0 a | |
| D | 0 a | ||
| Scarification (removal of epicarp and mesocarp) + IAA, 0.5mM | E0 | L | 0 a |
| D | 0 a | ||
| E1 | L | 0 a | |
| D | 0 a | ||
| Scarification (removal of epicarp and mesocarp) +IAA, 1mM | E0 | L | 0 a |
| D | 0 a | ||
| E1 | L | 0 a | |
| D | 0 a | ||
| df= 47, F=231.60, P=0.000 | |||
E0= Seeds of current year, E1= Seeds collected one year before,
GA3= gibberlic acid. IAA= Indole acetic acid.
Different letters a, b, c, d, e, f, indicate comparison of means based on Tukey’s test (≤ 0.05).
Percentage germination in various effective treatments (E0= Seeds of current year, E1= Seeds collected one year before, L= alternate light and dark treatment, D = continuous dark treatment).

The fruit and seed of Myriophyllum spicatum have been variously described by different researchers and they were given to different names. Fruit has been given the name as nut and seed as nutlet (Orchard, 1986). But according to the present investigation, it cannot be called as nut because nut is a single seeded indehiscent fruit. Based on the present stereo and scanning electron microscopic investigations the fruit is a schizocarp with four deep longitudinal sutures from which it splits into four individual seeds called nutlets. The schizocarpic fruits are intermediate between dehiscent and indehiscent fruits. In Myriophyllum spicatum the fruit instead of dehiscing rather splits into a number of segments each containing one or more seeds. Our observations are strongly supported by the findings of Aiken (1981).
During the present investigation, it was observed that Myriophyllum spicatum produces a copious amount of seeds and the seed set percentage ranged from 70.98% to 77.91% across different populations. The seed set depends upon various factors, including inflorescence size, position of inflorescence with respect to water surface, meiotic behavior and habitat. The standing water populations possess large inflorescences, which are always above the water surface and have more than thirty flowers per inflorescence. The pollen ovule ratio is high and the stigmas remain receptive for long duration along the whole length of the spike. All these features including normal meiotic behavior favor effective pollination and in turn high seed set. The small amount of variation observed in seed set between different standing water population’s viz. Dal Lake, Mansbal Lake and Hygam wetland may be due to the eutrophic environment of the Dal Lake if compared to Mansbal Lake and Hygam wetland. This is supported by the work of Madsen &Boylen (1989) on Myriophyllum seed ecology from an oligotrophic and eutrophic Lake. The plants inhibiting running waters did not produce any seeds. Probably the spikes in such populations do not withstand the pressure of running waters and pollen grains are washed away without causing effective pollination. Seed germination in Myriophyllum spicatum was found to be significantly influenced by various factors such as physical and chemical treatments, age of seeds and light conditions. The seeds in which seed coat was not removed (intact seeds) did not germinate even when treated with different concentrations of GA3, IAA and also after removal of epicarp and mesocarp. The study of Patten (1955), that shows that partial removal of stony endocarp of the Myriophyllum spicatum seeds is effective in causing seed germination, is in close agreement with the present study, however, the removal of epicarp and mesocarp causes seed germination is not in agreement with the present study.
The cold stratification treatment to some extent improved seed germination. This confirms that dormancy is due to the mechanical barrier of the seed coat, mainly because of hard stony endocarp of the seed. Low temperature of the seeds causes rupturing of the seed coat leading to breakdown in their dormancy (Patten, 1955). Similar results have been obtained by Teltscherova & Hejny (1973); Baskin & Baskin (1998) and Hay et al.(2008) in different species of Potamogeton. Under natural conditions, the freezing winter temperatures of the Kashmir valley may have promoter effect on seed germination. Cold stratification is an effective way of alleviating dormancy in many species especially those from temperate regions (Baskin & Baskin, 1998; 2004) which ensures that germination does not occur until spring commences when water temperature is more favorable to the vegetative growth of aquatic plants (Hay et al., 2008). The seed age has a significant effect on its germination. The present data confirms that M. spicatum seeds do survive desiccation. The freshly collected seeds as well as those dry stored up to one year germinate provided the seed coat is cut. Percent germination, however, declines with the aging of the seed. Similar results were obtained by Hay et al. (2000, 2008) in the case of Potamogeton. These observations indicate that no after ripening period is needed by the embryo of M. spicatum and the seeds are an important means of propagation in the habitats where water dries up for some period during the year. These observations are in agreement with the work of Kadano (1984), who observed that Potamogeton distinctus overcomes the long lasting drought in irrigation reservoirs under warm temperature conditions by the formation of seeds.
The seed germination varied significantly under continuous dark and alternate light and dark conditions. The percentage germination was found to be more in alternate light and dark conditions if compared to the seeds Kept in continuous dark conditions in all the treatments. These observations are in agreement with the results of Coble & Vance (1987). However, for aquatic plants effects of quality of light are less clear, although some quantitative effects are known (Spencer et al., 1971; Sharma & Gopal, 1978). The seeds did not germinate in trays provided with sediment and water. The reason for this is the hard seed coat that restricts the water absorption and low temperature needed by the seeds for their germination, causing prolonged seed dormancy.
Myriophyllum spicatum produces a significant amount of seeds in standing water bodies of Kashmir. However, the seed germination of the species is influenced by various factors such as light, chilling, age of seedsand even the under sediment environment of these water bodies. The successful germination of seeds due to various physical and chemical treatments under lab conditions is an indication of the fact that some part of the species population might also be contributed by sexual reproduction (asexual reproduction being the dominant wayof reproduction) through seed germination in nature.
The second author is highly thankful to CSIR, New Delhi, for providing financial assistance in terms of JRF Fellowship during this work. We are also thankful to the Director of USIC for providing SEM facilities.
1. Aiken SG. A conspectus of Myriophyllum (Haloragaceae) in North America. Brittonia. 1981;33:57-69.
11. Figuerola J, Green AJ. How frequent is external transport of seeds and invertebrate eggs by waterbirds. A study in Donana, SW Spain. Archives of Hydrobiology. 2002;155:557-565. Hartleb CF, Madsen JD, Boylen CW. Environmental factors affecting seed germination in Myriophyllum spicatum L. Aquatic Botany. 1993;45:15-25.
Note : All material which is updated in JATS TECH content is considered to be as sample.